what would make a trait more likely to be passed on to later generations
What do phylogenetic trees illustrate? These diagrams don't merely organize cognition of biodiversity - they also show us that living species are the summation of their evolutionary history.
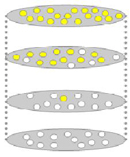
Imagine that a mutation occurred in an individual belonging to a lineage on a phylogenetic tree, and that this mutation changed some visible aspect of the organism that carried it. For example, suppose that the organism was a angiosperm, and that the mutation changed the establish's flower color allele then that the plant's blossoms were yellow rather than white. This mutant allele has the potential to either be lost from the population or to increase in frequency over many generations (Figure 1). The latter case is more probable when the mutant allele allows its carriers to produce greater numbers of offspring—in other words, the allele is more probable to increase in frequency when it confers higher fettle than the ancestral allele. In this case, natural selection volition favor fixation of the mutant allele.
No matter whether fixation of the derived allele occurs as a issue of pick or every bit a upshot of genetic drift, this fixation corresponds to extinction of the ancestral allele. Moreover, with the loss of the bequeathed allele, the population volition have evolved at the phenotypic level—specifically, the population that was formerly fixed for the possession of white flowers will now be made up of only yellow-flowered individuals. Considering the time information technology takes for a mutant allele to ascend and become fixed is generally very curt compared to the time between successive lineage-splitting events, it is usually safe to ignore the brief period when both the derived and ancestral alleles coexisted in the population, and to instead imagine that the derived trait arose in an evolutionary instant on some internal branch of the phylogenetic tree (Effigy ii).
Trait evolution is non anticipated. Nevertheless, once a lineage becomes stock-still for a derived trait, descendants of that lineage volition all have the derived trait unless there is a subsequent evolutionary change to a new trait (which might even resemble the original bequeathed trait). Thus, assuming there is not a subsequent change in flower color, all lineages descended from the original population that was fixed for yellow flowers will also have yellowish flowers.

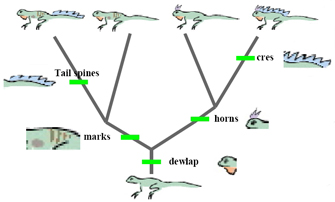
Of course, as phylogenetic trees branch, unlike lineages accumulate different traits. Figure 3 illustrates this idea using a clade that contains 4 cadger species. Y'all tin run into that the differences among these 4 species are the result of changes that occurred after the grouping began to diversify. In fact, if yous were told which traits evolved on which branches, you could precisely predict which traits each living species would have. Conversely, if given the features of each living species, you lot could explain the variation between the species by invoking just four events of trait evolution.
Every bit this example illustrates, descendants of an ancestral lineage tend to share common traits, and the presence of these characteristics allows scientists to infer the topology of phylogenetic trees. Although phylogenetic inference is a rich and complex field, information technology is based on the simple idea that every bit long every bit traits (or characters) change their state reasonably rarely (compared to the rate of lineage branching), then the distribution of traits among species provides evidence of how recently these species concluding shared a mutual antecedent.
One profound implication of this way of thinking about trait evolution is the idea that living species are the summation of their evolutionary history. In other words, if you lot list all the traits of a living species, you tin assume that each trait arose on an ancestral branch somewhere in that species' history. While some traits, such every bit the presence of a distinct cellular nucleus, evolved in the ancient by and are shared past many organisms, others, like spoken language, arose much more recently (in this case, very recently, as speech communication is a uniquely human trait). Thus, understanding the evolutionary history of living species amounts to agreement where on the tree of life these species' distinctive traits arose. Moreover, phylogenetic copse serve equally extremely powerful tools for organizing this knowledge of biological variety.
Phylogenetic Trees and Relatedness
In biology, the concept of relatedness is defined in terms of recency to a common ancestor. Equally a upshot, the question "Is species A more closely related to species B or to species C?" can be answered by request whether species A shares a more recent common ancestor with species B or with species C. To help clarify this logic, think almost the relationships within human families. The most recent common ancestors of both you and your siblings are your parents; the most contempo common ancestors of you lot and your first cousins are your grandparents; and the virtually recent common ancestors of you and your 2nd cousins are your great-grandparents. Note that your parents are situated one generation ago, your grandparents are situated two generations ago, and your swell-grandparents are situated three generations agone. This arrangement of ancestors explains why you lot are more closely related to your siblings than your cousins, and why you are more closely related to your first cousins than your second cousins.
Because evolutionary trees depict common beginnings, they also contain data on the degree of relatedness of the terminal nodes. For example, Effigy 4 can be used to determine whether a lizard is more closely related to a salamander or to a man. The first step in answering this question is to trace downward from the appropriate tips of the tree to find the most recent mutual antecedent of lizards and salamanders (marked x in the diagram). The next step is to use a similar process to locate the most recent common ancestor of lizards and humans (marked y in the diagram). Once you have done this, you lot can clearly encounter that node y is a descendant of node x, as indicated by the fact that you lot must laissez passer through node x to get from the root of the tree to node y. This tells you that lizards are more than closely related to humans than they are to salamanders.
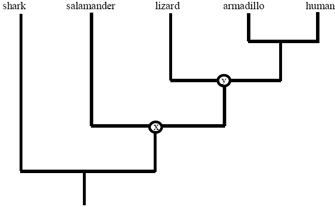
Many people are surprised by this conclusion regarding lizards, humans, and salamanders. This determination makes sense, notwithstanding, because there is a lineage (specifically, the internal branch betwixt ten and y) that is ancestral to both lizards and humans only not to salamanders. Any trait that evolved on this lineage will therefore tend to occur in lizards and humans only not salamanders. Ane such trait is the amnion, a protective layer that surrounds the embryo and originally served to allow eggs to mature away from h2o.
When it comes to this conclusion, one frequent source of confusion is the fact that salamanders and lizards look somewhat akin. However, while it is true that closely related organisms often take similar appearances, this is not e'er the case. The main reason for this observation is that morphological evolution tin occur at different rates on different branches of a phylogeny. In the example from Figure 4, the similarities of salamanders and lizards (e.g., sprawling gait and lack of fur) are features that trace back to the common ancestor of lizards, salamanders, and mammals. Among the land vertebrates, these are ancestral traits, or plesiomorphies. While the lizard and salamander lineages both retained these bequeathed traits, the mammalian lineage underwent dramatic evolutionary divergence, evolving an cock gait, fur, mammary glands, and many other features. These evolutionarily derived features, or apomorphies, are shared past all mammals merely are not establish in other living vertebrates. Withal, the presence of these uniquely mammalian features does not alter the fact that lizards are more closely related to mammals than to salamanders. Recollect, relatedness is nearly descent, not similarity. This should non be surprising. Subsequently all, you would notwithstanding be more than closely related to your first cousin than to your second cousin fifty-fifty if you happened to look more like to the latter!
"Tree Thinking": The Egalitarian View of Diversity

One of the most profound discoveries of evolutionary biology is the fact that all living species are continued through descent from a common ancestor. Thus, there is an underlying unity to life. At the same time, however, there is tremendous diversity in the living earth, which is the issue of the aggregating of unlike traits in different organisms. Thankfully, the tree metaphor not only offers a way to continue rail of the features of dissimilar organisms, merely information technology also provides guidance in how to conceptualize the broad sweep of biological diversity.
Since the time of the aboriginal Greeks, the prevailing guide to thinking about nature was the so-chosen "ladder of life," also known equally scala naturae. This idea imagines that living species correspond various degrees of perfection, with humans as the "most perfect" species and therefore at the top of the ladder (see, for case, Figure 5). Although the ladder of life idea was central to the evolutionary theories advanced by many of his predecessors, Charles Darwin largely rejected the ladder concept in favor of a tree; indeed, the simply figure in Darwin's Origin of Species was a tree, reproduced hither as Figure 6. Darwin'due south view of the evolutionary tree is beautifully expressed in the following quote:
The affinities of all the beings of the same class have sometimes been represented by a great tree. I believe this simile largely speaks the truth... The green and budding twigs may represent existing species; and those produced during sometime years may represent the long succession of extinct species . . . [T]he dandy Tree of Life . . . covers the earth with e'er-branching and cute ramifications.(Darwin 131–132)
In the years following Darwin's work, biologists formally rejected the ladder of life in favor of the tree concept. Nonetheless, many current discussions of evolution, especially in the general media, retain vestiges of the ladder view. This is often betrayed by the perception that some organisms are "advanced," whereas others are "archaic." Unfortunately, this ladder-based way of thinking about multifariousness is not but inefficient, but also positively misleading.
Why is this the case? For one, "ladder thinking" leads to statements that incorrectly imply that ane living species or group is ancestral to some other; examples of such statements include "tetrapods (state vertebrates) evolved from fish" or "humans evolved from monkeys." Only, is the goldfish an ancestor of any living tetrapod? And is the howler monkey actually an ancestor of you or any other human? Clearly, the answer to both of these questions is no. Why, then, does a statement like "tetrapods evolved from fish" seem reasonable at first glance?
"Tetrapods evolved from fish" might be intended to imply that the final common ancestor of a fish clade and a tetrapod clade was itself a fish. However, this is not strictly truthful, considering while the last common ancestor of both clades may have had more than obvious physical similarities to living fish than to living tetrapods, it was not identical to any living organism (fishlike or otherwise). Both lineages—the one leading to living fish (e.k., goldfish) and the one leading to living tetrapods—take been evolving independently for hundreds of millions of years, and during that time, evolution has not stood still on either the fish or tetrapod branch. Over this period, all aspects of fish physiology and the fish genome take changed, though mayhap in ways that are non obvious to the human eye. Thus, it is non authentic to say that the common ancestor of both fish and tetrapods was a fish. The all-time yous could do would exist to say that the mutual ancestor had a trunk class and environmental that were more similar to that of living fish than to that of living tetrapods.
Another problem with ladder thinking is that even with such clarifications, it is still like shooting fish in a barrel to brand errors of reasoning. For example, suppose you are told that goldfish have torso outgrowths (in this case, fins) with cartilaginous structures called rays. You are also told that tetrapods lack rays in their trunk outgrowths (limbs). If you took a progressive view, you might assume that tetrapods lost their rays during the form of evolution. In reality, nevertheless, the common ancestor of both tetrapods and goldfish lacked rays; thus, rays evolved forth the lineage that leads to goldfish. In this case, if you had assumed that the antecedent species had rays, ladder thinking would take led you lot astray. In order to avoid such mistakes, information technology is best not to make statements such as "tetrapods descended from fish," or at least to do so with the clear understanding that "fish" is referring only to body form and ecology and not to any other features of living fish species.
Tree thinking teaches u.s.a. that all living organisms are every bit distant in fourth dimension from the root of the tree of life and therefore all are equally advanced. Thus, in the eyes of evolution, a human and a bacterium are as derived. Although one of these organisms is certainly more morphologically circuitous than the other, both organisms are remarkable in that they are the product of parents that successfully and repeatedly gave rise to offspring over an unimaginably long fourth dimension bridge (at least 3 billion years).
This egalitarian view of life may seem hard to swallow. However, before you reject this idea, consider how the world might look if you were a ladder-thinking bacterium. If that were the case, you would certainly exist struck by all the astonishing molecular adaptations that your ancestors had accumulated to make you and your kin and then successful. Yous would probably indicate to a human and note that within its body, there are more bacterial cells than human cells, thereby proving the superiority of bacteria over lumbering eukaryotes. You lot would probable consider bacteria to be the pinnacle of cosmos and the rest of the planet's organisms to be evolutionary rejects. On the other hand, if you were a tree-thinking bacterium, your view of life's tapestry would be just like that of the tree-thinking human—in this instance, you would appreciate that all living things are equally amazing products of over 3 billion years of evolution. Thus, tree-thinking non only provides important practical tools for organizing knowledge of biodiversity and for reconstructing evolutionary history, just it too provides a clear and unbiased metaphor for evolution at large.
References and Recommended Reading
Avise, J. C. Evolutionary Pathways in Nature: A Phylogenetic Approach (Cambridge Academy Printing, Cambridge, UK, 2006)
Baum, D. A., DeWitt Smith, S., & Donovan, S. S. The tree thinking challenge. Science 310, 979–980 (2005)
Baum, D. A., & Offner, S. Phylogenies and tree thinking. American Biological science Teacher 70, 222–229 (2008)
Darwin, C. On the Origin of Species by Means of Natural Option, or the Preservation of Favoured Races in the Struggle for Life (John Murray, London, 1859)
Dawkins, R. The Ancestor's Tale: A Pilgrimage to the Dawn of Life (Houghton Mifflin, New York, 2004)
Delsuc, F., et al. Phylogenomics and the reconstruction of the tree of life. Nature Reviews Genetics 6, 361–375 (2005) doi:10.1038/nrg1603 (link to commodity)
O'Hara, R. J. Homage to Clio: Toward an historical philosophy for evolutionary biology. Systematic Zoology 37, 142–155 (1988)
O'Hara, R. J. Population thinking and tree thinking in systematics. Zoologica Scripta 26, 323–329 (1997)
Maddison, W. P., & Maddison, D. R. MacClade: Assay of Phylogeny and Character Development (Sinauer Associates, Sunderland, MA, 1992)
Tree Thinking Group. Tree Thinking Grouping homepage, (2004)
johnsonclithapping.blogspot.com
Source: http://www.nature.com/scitable/topicpage/trait-evolution-on-a-phylogenetic-tree-relatedness-41936
0 Response to "what would make a trait more likely to be passed on to later generations"
Post a Comment